

From the Townsend Letter |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
How Can We Cure NO/ONOO− Cycle Diseases? |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
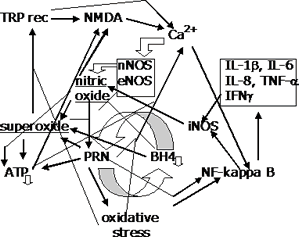
Abstract The basic concept of the NO/ONOO− vicious cycle mechanism is simple. It is that various short-term stressors can initiate this cycle which, like all vicious cycles, propagates itself over time. The cycle then, depending on where it is located in the body, causes various chronic diseases. But in order to treat chronic diseases caused by the NO/ONOO− cycle and hopefully cure them, one needs to understand the details of the cycle mechanism. And that is where things becomes much more complex. Probably the most important part of the cycle is what I have called the central couplet, the reciprocal relationship between elevated peroxynitrite (abbreviated PRN) and the depletion of a compound called tetrahydrobiopterin (BH4) (see Figure 1 center to below center). Peroxynitrite oxidizes BH4 at physiologically relevant concentrations leading to a BH4 depletion.9-11 BH4 is a cofactor in nitric oxide synthases, such that when these NOSs are missing BH4, they become uncoupled, producing superoxide in place of nitric oxide. In partial uncoupling, this superoxide can react in turn with the nitric oxide produced by adjacent coupled enzymes, leading to more peroxynitrite. Because the reaction between superoxide and nitric oxide is extraordinarily rapid, what is called diffusion controlled, the production of both molecules by adjacent enzymes may be particularly effective in raising peroxynitrite levels. Thus, although this partial uncoupling lowers nitric oxide production, it is expected to increase peroxynitrite production, the most central part of the wider NO/ONOO− cycle.1-3,11 The superoxide produced by such partial uncoupling has a special role, then, in producing peroxynitrite. It should be noted that the production of superoxide remote from the production of nitric oxide will be much less effective in raising peroxynitrite levels because there are high amounts of superoxide dismutase in cells and extracellular fluid, which destroys most of the superoxide before it travels very far from its site of synthesis. Types of Evidence Supporting the NO/ONOO− Cycle Mechanism
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

flavonoids, ecklonia cava extract, algal supplements |
chain breaking and other antioxidant activity |
Some may act as peroxynitrite scavengers. |
NMDA antagonists, other agents that indirectly lower NMDA activity; magnesium |
All act to lower excessive NMDA activity |
|
acetyl carnitine/carnitine, coenzyme Q10, low hyperbaric or normobaric oxygen |
Improved mitochondrial function |
Oxygen must be used with caution, particularly in severe cases of CFS/ME |
hydroxocobalamin form of vitamin B12 |
Reduced in vivo to a form that is a potent nitric oxide scavenger. |
Higher dosage (i.e., 5 to 10 mg) needed than is needed to treat a B12 deficiency; Typically used via IM injection, as an inhalant, or via nasal spray to obtain high blood levels; oral or sublingual should be useful but are clearly suboptimal because of limited absorption. |
high-dose folates |
Serves as precursor of 5-methyltetrahydrofolate (5-MTHF), a potent peroxynitrite scavenger. |
Unclear whether folic acid, folinic acid, 5-MTHF and/or other forms of folate should be used; folic and folinic acid tested in published trials. |
D-ribose, RNA, inosine |
All act to increase uric acid levels (peroxynitrite scavenger); all may act to help restore ATP pools. |
Published trial on D-ribose; trial currently in progress suggesting inosine can be helpful. |
IV high dose, buffered ascorbate |
Lowers both ends of central couplet (see below); may be particularly helpful agent. |
Discussed in detail below. |
sauna therapy |
Acts to increase BH4 availabiity; mechanism via increased synthesis of GTP cyclohydrolase I.13 |
Trials published on MCS, FM and CFS/ME; discussed further below. |
fish oil |
Established as anti-inflammatory agent. |
May also improve brain function. |
Most studies involved CFS/ME and/or FM; however studies with sauna therapy and IV ascorbate have been published with MCS patients.
It can be seen from Table 1 that agents lowering various aspects of the NO/ONOO− cycle are helpful in treatment of these three apparent NO/ONOO− cycle diseases. Specifically, agents that lower oxidative stress, lower peroxynitrite, improve mitochondrial function, lower NMDA activity, increase BH4 availability, or have anti-inflammatory activity all appear to be helpful in treatment. It is difficult to see how this could be the case unless the NO/ONOO− cycle or something very similar to it is the central cause of these multisystem diseases.
The evidence summarized in Table 1 also strongly suggests that the NO/ONOO− cycle makes very useful predictions in terms of therapy. Given the complexity of the cycle, as diagrammed in Figure 1, it seems likely that multiple agents lowering various aspects of the cycle will be most effective in producing clinical improvements in apparent NO/ONOO− cycle disease.
Multiagent Protocols and the Allergy Research Group Nutritional Support Protocol
I described the responses to five multiagent protocols developed by different researchers, one of which I had a role in developing, in Chapter 15 in my book.1 Each of these five involved 14 to 18 different agents or classes of agents that are predicted to downregulate one or more aspects of the NO/ONOO− cycle. Each of these apparently produces substantial improvements in many patients suffering from these multisystem diseases, although four of the five have only been tested on one disease. In contrast, Teitelbaum's protocol has been tested on both CFS/ME patients and FM patients with apparent positive results.14
More recently I have developed, in collaboration with the Allergy Research Group, an entirely over-the-counter protocol based on nutritional supplements chosen to downregulate the NO/ONOO− cycle. This nutritional support protocol is described elsewhere as well as on a page of my website (thetenthparadigm.org/arg.htm).2,3 It includes 22 agents chosen for their ability to downregulate various aspects of the NO/ONOO− cycle as well as other general nutrients.
The feedback I have gotten from clinical observations of physicians and others who have used it to treat their CFS/ME, FM, and/or MCS patients is that 80% to 85% of patients respond positively to it and that improvements are typically maintained if patients can avoid exposure to stressors that are predicted to upregulate the cycle.2,3 Even patients who have been ill for two decades or more often respond positively. It should be noted, however, that the extent of improvement tends to vary considerably, from responses described as miraculous to modest improvements. Furthermore, about 15% to 20% of patients do not improve. Patients with high levels of mercury in their bodies may react negatively to the protocol, presumably because mercury can be mobilized by the alpha-lipoic acid found in the protocol. One group who appear not to respond substantially, either positively or negatively, are the chronic Lyme disease patients.
Note: I must point out to the reader that I do have a conflict of interest here. I receive a small royalty from the Allergy Research Group for designing much of this nutritional support protocol.
Ingrid Franzon and colleagues in Sweden have run a small pilot study on the Allergy Research Group protocol with a group of nine CFS/ME patients (personal communication). She found, surprisingly for such a small group, statistically significant improvement in physical health measures within four weeks, with a further statistically significant improvement over another four weeks (personal communication; statistical analysis performed using a paired Student's t test).
In summarizing the last two sections of this article, there are four important points to be made: First, individual agents that downregulate specific parts of the NO/ONOO− cycle have often been reported in clinical studies to produce improvements in these multisystem-disease patients. Second, multiple parts of the cycle are implicated from these clinical studies as well as from other studies, producing strong confirmation that a mechanism like the NO/ONOO− cycle is the central etiologic mechanism of these diseases. Third, and not surprisingly, treatment protocols using multiple agents predicted to downregulate the cycle seem to be more effective than single agents. Fourth, these several multiagent protocols are not effective with patients who are repeatedly or continuously exposed to stressors that otherwise upregulate the NO/ONOO− cycle.
Clearly, these four points are very important and exciting, considering that the conventional wisdom has been that there is little that can be done to treat these illnesses.
My own view is that the Allergy Research Group nutritional support protocol is the most promising of these multiagent protocols, because it is relatively inexpensive; it is available over the counter in the US, Canada, and much of Europe; and it apparently achieves good responses despite the limitations inherent in over-the-counter approaches.
The NO/ONOO− cycle etiology is best documented for CFS/ME, MCS, FM, and PTSD, as well as for Gulf War syndrome/illness, which appears to be a combination of the four.1-7 However, there is also at least a superficial case to be made that 14 additional diseases, including the classic neurodegenerative diseases asthma, multiple sclerosis, tinnitus, and autism, appear to also be caused by the NO/ONOO− cycle.1 The cases that have been made for each of these 14 are frankly relatively superficial, except for tinnitus and postradiation syndrome, where more extensive cases have been made.1,15,16 Thus this approach to the treatment of chronic disease may not be limited to such diseases as CFS/ME, FM, MCS, and PTSD but have vastly broader implications.
How Can We Start Getting Substantial Numbers of Cures?
The use of multiple agent protocols where individual agents act to lower the NO/ONOO− cycle is an exciting and promising approach to treating these diseases. However, based on published and (to the extent I have access to it) unpublished evidence, none of these protocols produces any substantial numbers of cures. If we understand the NO/ONOO− cycle mechanism sufficiently and if we are effectively downregulating it, we should start seeing substantial numbers of cures. Why has this not happened?
My own view, is that the central couplet is insufficiently downregulated in these protocols. The main argument explored in this article is that by more effectively lowering this central couplet mechanism, we may be able to extend these multiple agent protocols to obtain substantial numbers of cures. As described, the central couplet is the reciprocal relationship between peroxynitrite elevation on the one hand and BH4 depletion on the other. We need to focus, then, on agents that lower peroxynitrite and its products at one end of the central couplet, and agents that raise BH4 availabliity on the other end.
There are at least ten available agents that are predicted to substantially lower the central couplet, summarized in Table 2; and we will explore each of them one at a time.
Table 2: Agents/Classes of Agents Predicted to Sustantially Lower the Central Couplet
Agent Dosage Presumed Mechanism(S)
IV buffered ascorbate |
7–50 g, |
1. Acts as peroxynitrite scavenger; 2. reduces B back to BH4, helping restore BH4 levels; 3. the very high levels obtained by IV treatment can lead to increased levels of hydrogen peroxide, leading to induction of GTP cyclohydrolase I, thus leading to increased de novo synthesis of BH4. |
oral ascorbate |
circa 2–3 g, repeated daily |
Blood levels obtained are substantially lower than for IV treatment, above. However such levels may be adequate to trigger the first two mechanisms outlined immediately above. |
sauna therapy |
repeated |
Induces GTP cyclohydrolase I, leading to increased de novo synthesis of BH4. |
reduced glutathione, liposomal, time release, nasal spray, IV or inhalant |
150–500 mg per day |
Reduces BH2 back to BH4, thus helping restore normal BH4 levels and lowering the partial uncoupling of the nitric oxide synthases; some, particularly those with asthma-type symptoms, may have some difficulty tolerating this treatment, depending on dosage regimen. |
Inosine, RNA, or D-ribose |
varies |
Each of these has the capability of producing two responses: restoration of adenine nucleotide pools and increased uric acid levels in blood. The latter will lead to lowered levels of peroxynitrite breakdown products, NO2 radical and carbonate radical. Each of these agents have drawbacks (see text). |
5-methyl tetrahydro- |
300 µg/day for 5-MTHF, higher doses for precursors |
Acts as a potent peroxynitrite scavenger and will therefore help also restore BH4 pools; high dose folic or folinic acid will act to help raise 5-MTHF pools. 5-MTHF pools are depleted in CFS/ME, presumably due to peroxynitrite mediated oxidation. |
tetrahydro- |
circa 5 mg or less, oral daily |
Helps restore BH4 pools; also acts as peroxynitrite scavenger. This would be an off-label use of BH4. |
vasoactive intestinal peptide (VIP) |
IV or inhalant |
Induces GTP cyclohydrolase I, leading to increased de novo synthesis of BH4; this would be an off-label use. |
flavonoids, ellagic acid, other phenolic antioxidants |
??, oral |
Probably act to scavenge peroxynitrite and breakdown products and may also act more directly to help restore BH4; dosage and optimal sources are unclear. |
hydroxocobalamin |
IM injection, nasal spray or inhalant |
Acts in the reduced form (cobalt II) as a potent nitric oxide scavenger; this will indirectly lower peroxynitrite because of the role of nitric oxide as a peroxynitrite scavenger. |
Taken from the author's website with permission.
General Strategy and Relationship to Lowering Hypertension
The general strategy is that an effort to lower the central couplet will be made, as a second phase to a treatment in which the first phase is a wide-ranging protocol lowering other aspects of the cycle together with avoidance of stressors that will otherwise upregulate the cycle. The Allergy Research Group nutritional support protocol for doing this is described on my website (thetenthpardigm.org/arg/htm), as well as elsewhere.2,3 The overall concept is that by lowering various aspects of the cycle and then focusing on lowering the central couplet, we should start seeing some cures of NO/ONOO− cycle diseases. Let me remind the reader, that I am a PhD, not an MD, and nothing I say or write should be viewed as medical advice.
A second part of this general strategy is that by using relatively high doses of agents that act collectively to lower both ends of this central couplet, one may get responses that may go up as much as the square of the dose of these combinations of agents. Relatively high doses of agents that are acting at the same time to lower both sides of the couplet may be most effective. This may be expected because this central couplet is just that, a couplet wherein lowering peroxynitrite will increase the availability of BH4, and independently increasing the availability of BH4 will lower peroxynitrite and its products. Therefore, doing both of these simultaneously can have a major impact in lowering this central couplet.
The effectiveness of agents in accomplishing this task may be judged to some extent by their ability to lower hypertension. Hypertension is thought to be caused by shifting the ratio of nitric oxide to peroxynitrite, towards excessive peroxynitrite, something that is produced by the action of the central couplet. This role in hypertension is a consequence of the following: whereas nitric oxide is a vasodilator, peroxynitrite is a vasoconstrictor, acting in part by raising the levels of isoprostanes, which are potent vasoconstrictors. For example, vasopressin II acts to produce hypertension by inducing higher levels of NADPH oxidase, an enzyme whose activity produces superoxide.17 The reaction of superoxide with nitric oxide will produce peroxynitrite and thus turn on the central couplet. Depletion of BH4 levels has been shown to have an important role in causing hypertension.18-20
Treatments that lower hypertension may be suggested to be effective agents in lowering the central couplet. However, because hypertension occurs outside the central nervous system but some NO/ONOO− cycle diseases may be localized to a great extent in the brain, agents that fail to traverse the blood–brain barrier may act on hypertension but may not effectively lower central nervous system–located NO/ONOO− cycle diseases. It is important, therefore, to keep this restriction in mind, because it may limit the prediction that treatments that lower hypertension will work to produce improvements in NO/ONOO− cycle diseases.
Let us discuss the apparent mechanisms of action of the 10 agents discussed in Table 2.
IV Ascorbate
Intravenous (IV) ascorbate (vitamin C) can produce levels of ascorbate in the blood of 100 times or more the upper level of "normal."21-24 By doing so, it may produce effects vastly greater than one will get from normal pools sizes of ascorbate. IV ascorbate, typically using 7 to 50 g of buffered ascorbate, has been successfully used to treat MCS or CFS/ME patients; and, in addition, I am aware of a number of physicians who have reported successfully treating these patients with such doses of buffered IV ascorbate.25-28 Such IV ascorbate treatments appear to be well tolerated, even at doses roughly 4 times the highest doses suggested here, except possibly when two contraindications are present (see below).21-24,29
There are three effects of ascorbate that may be expected to occur in response to such high levels:
- Ascorbate is a scavenger of peroxynitrite and its breakdown products, but has only modest scavenging activity at normal ascorbate blood levels.30-32 It will be expected to have much greater scavenging activity with levels many times the normal upper level.
- When peroxynitrite oxidizes BH4, the initial product is B, the one electron oxidation product. B can be reduced back to BH4 by ascorbate, which is, of course, a reducing agent.30,32 However, B is itself unstable and will probably therefore require high levels of ascorbate to efficiently produce such reduction.9,10,30
- The very high levels of ascorbate produced by such IV treatment produces hydrogen peroxide via ascobate oxidation and concomitant reduction of molecular oxygen.21-24,33,34 Hydrogen peroxide is known to be able to induce the enzyme GTP cyclohydrolase I, the first and rate-limiting enzyme in the de novo pathway to synthesize BH4.35-37 It follows that IV ascorbate may be expected to increase the availability of BH4 by this mechanism, as well as by the preceding one.
It follows that IV ascorbate may be able to favorably affect both sides of the central couplet, lowering peroxynitrite and its products and also, via two distinct mechanisms, increasing availability of BH4. This set of three mechanisms collectively produces a rationale for the use of IV ascorbate in the treatment of these multisystem illnesses. To my knowledge, there has been no previous rationale for such treatment, despite its reported effectiveness.
It will probably be important to determine that patients to be treated with such IV ascorbate do not have highly elevated levels of free iron, to avoid triggering extensive Fenton chemistry with the ascorbate treatment. Typically, this means that serum iron binding capacity should be no more than the upper limit of "normal"; that is, no more than 55% saturated.
In addition, those with a genetic glucose-6-phosphate dehydrogenase (G6PD) deficiency are susceptible to hemolysis caused by IV ascorbate because they are less able to detoxify the consequent hydrogen peroxide, so that treating such patients with IV ascorbate is contraindicated.38 Patients should therefore be tested for possible G6PD deficiency and for elevated free iron, and only those lacking both of these contraindications should be treated with high-dose IV ascorbate.
IV ascorbate used in such treatment should be buffered to the physiological pH of the blood (7.4) to avoid shifting the pH. Such buffering particularly important in those with kidney dysfunction who are less able to regulate the pH of the blood.
With the exception of cancer treatment, where IV ascorbate is thought to act mainly via increased production of hydrogen peroxide, there has been no widely applicable rationale for its reported effectiveness in the treatment of other diseases. The mechanisms described in this section are important, therefore, in providing such a rationale, one that makes important predictions about how IV ascorbate treatment may be useful and what strategies can maximize its efficacy.
Oral Ascorbate
Oral ascorbate can yield levels typically circa three times the upper range of normal with doses of 2 to 3 g. Such doses and somewhat lower doses are reported to lower hypertension, suggesting that they may be able to decrease the central couplet.18,39-41 Typically, such high blood levels are only maintained for relatively short periods of time, on the order of 4 hours.22 Although 2 to 3 g of oral ascorbate lead to absorption over 3 to 4 hours, there is also rapid excretion of high blood levels of ascorbate; that is, of levels well in excess of normal.22 The levels of ascorbate produced by 2 to 3 g or higher doses of oral ascorbate may be expected to trigger substantial peroxynitrite scavenging, as well as some chemical reduction of B to BH4, but not any substantial hydrogen peroxide–induced increased levels of GTP cyclohydrolase I (see previous section).30-32
It follows that such doses of oral ascorbate may be expected to act to lower the central couplet, although they will be less active in so doing than the much higher IV doses.
Sauna Therapy
Sauna therapy has been reported to be helpful in the treatment of MCS, FM, and CFS/ME, as well as with other diseases characterized by BH4 depletion. 42-50 Sauna therapy is thought to act via two distinct mechanisms to induce higher levels of GTP cyclohydrolase I and thus increased availability of BH4.50 Substantial increased availability probably only occurs after repeated sauna treatment.
In terms of strategy, therefore, it seems likely that sauna therapy could be useful in trying to cure these diseases, as follows: after several sauna treatments, subsequent sauna treatments should be accompanied by treatments with one or more agents that scavenge peroxynitrite and possibly also one or more agents that help reduce previously oxidized biopterin forms, such as B and/or BH2, to BH4.
Reduced Glutathione
BH4 oxidation by peroxynitrite produces initially B, much of which is rapidly oxidized further to BH2, the two electron oxidation products. BH2 can be reduced back to BH4 by reduced glutathione and other thiol compounds.30 Therefore, raising reduced glutathione levels may be useful in restoring BH4 availability.
Oral glutathione typically gets degraded in the GI tract, but a number of approaches can be used to try to increase it. These include using oral liposomal or possibly time-release oral glutathione, or reduced glutathione via nasal spray, IV, or nebulized inhalant. Reduced glutathione has, of course, several other antioxidant properties that should make it useful in the treatment of NO/ONOO− cycle diseases, so clearly its actions are not specific to lowering of the central couplet.
There is one complication to using reduced glutathione in those who have asthma-type responses: they report that reduced glutathione treatment may trigger asthma attacks. I think that this is probably due to the action of reduced thiols in activating some of the transfer receptor potential (TRP) receptors, including TRPA1. In any case, it can cause problems. This problem is probably most substantial when using inhaled nebulized glutathione, but other treatment modalities may occasionally cause such reactions. Nevertheless, I am aware of reports that reduced glutathione treatment can be very helpful in the treatment of NO/ONOO− cycle diseases, so it should be considered as a therapeutic agent.
Inosine, RNA, or D-Ribose
I have lumped these three agents together because each of them is expected to produce two specific favorable responses. One of these two responses acts to lower the central couplet. Let's consider the response unrelated to the central couplet first, and then the one that lowers the couplet.
Each of these three agents will raise levels of purine nucleotides in the body, including the adenine nucleotides that comprise ATP and others (ADP and AMP) that can act as precursors for ATP. ATP is, of course the "energy currency" in the body, and its levels will be depleted whenever there is mitochondrial dysfunction. Because mitochondrial dysfunction is part of the NO/ONOO− cycle, this will occur in cycle diseases. When it is sufficiently severe, it will lead to accumulation of fairly large amounts of AMP, which will be degraded further, lowering the levels of all of these adenine nucleotides (that is, ATP+ADP+AMP). This causes a longer-term problem, because when and if there is an improvement in mitochondrial function, the lowered adenine nucleotide levels mean that the cell has a problem is producing normal ATP pools, even when the mitochondria are otherwise capable of doing so. Each of these three agents will allow the production of increased adenine nucleotides, potentially leading in turn to increased ATP. This is the interpretation that has been given to the improvements reported for D-ribose treatment of both CFS/ME and FM, and it may be partly responsible for that improvement.51
However, there is a second response to all three of these agents that will directly lower the central couplet. Increased purine nucleoside and nucleotide pools will subsequently produce increased purine degradation, the end product of which is uric acid, an important scavenger of peroxynitrite and its breakdown products in humans.31,52 Of course, by lowering peroxynitrite and its oxidant products, uric acid will lower the central couplet.
Uric acid levels in the blood are often about 4 to 5 times those of ascorbate, although there is quite a bit of variation around those figures. However the effectiveness of uric acid in scavenging peroxynitrite and its products, per mole, is roughly similar to that of ascorbate.31 Consequently, even though it is possible to raise ascorbate levels by much higher percentages than uric acid levels in vivo, it seems likely that raising uric acid levels will produce a substantial effect on peroxynitrite-mediated oxidations in vivo and therefore should be considered well worth pursuing in lowering the central couplet.
Uric acid has a half-life of about 20 hours in humans, so it should not take very long to increase its levels by increasing the availability of purine-containing compounds in the body, such that when an increase in purine degradation is obtained, it will be sustained substantially longer than any high-level ascorbate elevation.53 Consequently, it makes sense to consider each of these three supplements – inosine, RNA, and D-ribose – as possible agents to raise uric acid levels.
While each is expected to be helpful in two ways, one of which lowers the central couplet, each of these three agents also has a possibly problematic feature:
D-ribose is a potent glycating agent, being approximately 50 times more active in glycation than is D-glucose (the normal sugar in the blood), with substantial possible physiological effects of such D-ribose mediated glycation.54-56 Protein glycation is associated with aging and produces dysfunction of many glycated proteins.
The commercial source of RNA is yeast, and some sufferers of these diseases have yeast allergies and so may have difficulty in tolerating RNA.
Inosine is in general a well-tolerated supplement.57 However, it can stimulate the activation of mast cells, and people with these illnesses often have problems with excessive mast cell activation. Inosine is known to act to stimulate mast cell activation via the adenosine A(3) receptor.58
Of these issues, the one that concerns me the most is the glycation via D-ribose, although I know that Dr. Jacob Teitelbaum, whom I respect greatly, disagrees with me on this.
People with these diseases tend to be low in uric acid, presumably because of the oxidation of uric acid by peroxynitrite and its breakdown products. Because of the important role of uric acid in lowering peroxynitrite-mediated damage, it seems likely that raising uric acid levels may be an important approach to lowering the central couplet. One does need to be careful not to raise uric acid levels too much, because excessive levels can cause gout. In normal people, this is not a problem, because uric acid excretion greatly increases as blood levels exceed normal levels; but it may be a concern in those who are susceptible to gout, where the excretion mechanism may not function properly.
A second, related issue is that very high uric acid levels may produce hypertension, and while direct measurements suggest that uric acid lowers nitric oxide synthase uncoupling, rather than raising it, this also suggests that we should limit the rise in uric acid levels in these treatments.
With these two caveats in mind, a substantial rise in uric acid levels into the mid-to upper-normal range may be very helpful to people suffering from NO/ONOO− cycle diseases.
5-Methyltetrahydrofolate (5-MTHF)
It has been known for a number of years now that high-dose folic acid supplements can lower partial nitric oxide synthase uncoupling (this has been most studied with the eNOS nitric oxide synthase form), with much of this effect being due to increased availability of BH4.59-62 This response depends on the reduction of the folic acid by the enzyme dihydrofolate reductase, showing that a reduced form of folate probably has a role here. What has been unclear until recently is the reduced folate's mechanism of action.
It has been shown, however, that 5-methyltetrahydrofolate (5-MTHF) is an extremely potent peroxynitrite scavenger, so the probable mechanism of action is the lowering of peroxynitrite and its breakdown products.63,64 In other words, this is another situation in which the central couplet is involved, such that by lowering one end of the couplet (the peroxynitrite end), one also lowers the other end (increasing BH4). Another reduced folate, tetrahydrofolate, also acted as a peroxynitrite scavenger, although it was less active than was 5-MTHF.63
This action of 5-MTHF is also supported by its role in vivo and in vitro as an extremely active scavenger of singlet oxygen.65 Singlet oxygen is known to share chemical similarities to peroxynitrite because both molecules have very weak oxygen–oxygen bonds, so the similar scavenging of both molecules by 5-MTHF should not be surprising.
It has been shown that high-dose oral folic acid can lead to major increases in 5-MTHF. For example, Doshi et al. in their figure 5 showed that a single 5 mg folic acid supplement in humans led to roughly seven times the initial blood levels of 5-MTHF in 3 to 4 hours.66 They also showed that repeated daily 5 mg doses produced still higher 5-MTHF levels, roughly 15 times the initial levels, an effect that they attributed in part to an induction of the dihydrofolate reductase enzyme.
Jacobson et al. showed that levels of 5-MTHF in the sera from CFS patients were very low compared with normals and that other reduced folate pools were also depressed.67 I am aware of extensive unpublished data on CFS/ME patients, confirming these results. Gerwin reported that folate deficiency was one of the three most common systemic factors in myofascial pain syndrome, a condition closely linked to fibromyalgia.68 These studies strongly suggests that elevated peroxynitrite levels in CFS/ME and possibly other multisystem illnesses may produce a substantial loss of 5-MTHF, and that some of the products of 5-MTHF oxidation are lost to the folate pools, thus leading to an overall lowering of folates in the body. The lowering of 5-MTHF pools has also led in the unpublished data to a much more modest (circa 10% to 15%) lowering of S-adenosylmethionine levels.
It can be inferred from the studies discussed in this section that the reaction between 5-MTHF and peroxynitrite can have substantial impacts on both 5-MTHF levels and peroxynitrite-mediated responses in real physiological situations. With regard to the main focus of this article, raising the levels of 5-MTHF can significantly affect the central couplet by lowering the levels of peroxynitrite and its breakdown products. The practical question is whether this can be best accomplished by using high folic acid doses, which act as a precursor for 5-MTHF, or using 5-MTHF itself and/or other reduced folates that can serve as precursors of 5-MTHF, such as folinic acid. The answer is uncertain.
There are two important complications to this story. I have received information from two sources to the effect that using doses of 5-MTHF in substantial excess of 300 mcg leads to negative reactions in patients suffering from presumed NO/ONOO− cycle diseases. My guess is that this may be due to the toxicity of some of the oxidation products of peroxynitrite-mediated oxidation of 5-MTHF. If this interpretation is correct, it may be possible to increase the well-tolerated dose if one uses other agents that lower peroxynitrite at the same time.
The second complication is that there must be very rapid turnover of the methyl group on intracellular 5-MTHF. There are massive amounts of methylation going on in the body, and even though the great majority of that does not go through 5-MTHF, there still must be rapid turnover of the methyl group on 5-MTHF. It follows that the half-life of intracellular 5-MTHF is probably on the order of few seconds, and while the 5-MTHF can be regenerated after it acts as a methyl donor, the efficiency of that process is uncertain. Consequently, the effectiveness of an oral supplement of 5-MTHF on the scavenging of peroxynitrite may be expected to be greater in the extracellular space than it is intracellularly.
Folinic acid supplements were shown to produce major improvements in a group of CFS/ME patients.69 A number of other studies have reported major improvements in CFS/ME or FM patients with treatment protocols including high-dose folic acid or other folates, but it is difficult to determine the role of the folates themselves in such complex protocols.
Based on the compelling biochemistry, I think that folates, both folic acid and reduced folates, are among the most attractive agents in lowering the central couplet.
Tetrahydrobiopterin (BH4)
Perhaps the most obvious agent to use to lower the central couplet is BH4 itself, or alternatively precursors such as sepiapterin or biopterin. BH4 supplements have been reported to be helpful for the treatment of autism patients, and autism is one of the proposed NO/ONOO− cycle diseases.1,70-72 There are, however, some complications that need to be considered in using BH4 to lower the central couplet.
First, it is known that oral BH4 is largely oxidized and must therefore be reduced back to BH4 before it can function in target cells. Most of this reduction occurs intracellularly through enzymatic reduction. However, the rapid peroxidation of the BH4 leads to questions of whether this oxidation may produce peroxidative damage. For example, although Parkinson's disease is thought to involve BH4 depletion, an animal model study showed that high doses of BH4 produced Parkinson's-like symptoms and neuronal damage, providing some support for this view.73,74 In any case, it may be important to limit the dosage of BH4 if it is used directly to prevent any major consequences of BH4 peroxidation. It is possible that reducing agents such as high-dose ascorbate may minimize this peroxidation, and that using BH4 along with high-dose ascorbate may be helpful in constructing therapeutic strategies.
An alternative approach is to use precursors such as biopterin or sepiapterin as oral supplements to provide increased availability of BH4.
Vasoactive Intestinal Peptide (VIP)
VIP has been used by two physicians to treat CFS/ME patients or chemically sensitive patients (unpublished data), with apparently good responses in both. For example, Dr. William Rea has used VIP with his chemically sensitive patients with apparently good responses (personal communication). VIP is known to lower several parts of the NO/ONOO− cycle, and the most likely mechanism for this, in my view, is the reported role of VIP in inducing GTP cyclohydrolase I activity and consequently raising BH4 levels.75 This view is supported by the well-documented role of VIP in improving vasculature function. VIP is known to lower hypertension and vascular endothelial dysfunction, and both of these are caused in part by BH4 depletion.
Flavonoids, Ellagic Acid, and Other Phenolic Antioxidants
A number of flavonoids have been shown to act as scavengers of peroxynitrite, and also its precursor superoxide it has been suggested that they can be active in vivoin lowering peroxynitrite-mediated effects.76 Other phenolic antioxidants can also have important roles here, and perhaps one of the most important may be ellagic acid, which scavenges peroxynitrite.77 It is not clear to me which sources of these phenolics are the most likely to be useful here, but perhaps pomegranate extract, which contains substantial amounts of ellagic acid, and also several flavonoid-containing extracts that are reported to lower hypertension and improve vascular endothelial dysfunction.78-82 Ghosh and Scheepens list cocoa, wine, grape seed, berries, tea, tomatoes (polyphenolics and nonpolyphenolics), soy, hawthorn, and pomegranate as attractive possibilities for phenolic antioxidants that may lower hypertension and improve vascular endothelial dysfunction.80 Schmitt and Dirsch list cocoa, pomegranate, both green and black tea, olive oil, and soy among food sources. They also list ginkgo, hawthorn, and ginseng among herbal sources.81 Extracts of each of these should be considered as agents for possibly lowering the central couplet.
Hydroxocobalamin Form of Vitamin B12
Hydroxocobalamin has been used for over 70 years to decrease fatigue in people with chronic fatigue, long before CFS/ME was a well-defined illness. It was shown in a clinical trial of patients with a CFS/ME-like illness that 5 mg intramuscular (IM) injections twice a week produced statistically signficant improvements as compared with placebo.83 In this study, it was also shown that there was no correlation between initial B12 levels and response to hydroxocobalamin therapy, suggesting that the hydroxocobalamin was not acting primarily to allay a B12 deficiency. Lower doses of another form of B12 that were adequate to allay a possible B12 deficiency produce no clinical improvement, and other evidence also strongly suggests that high-dose hydroxocobalamin is not acting here to allay a B12 deficiency.84,85
Other uncontrolled studies have suggested that the hydroxocobalamin form of vitamin B12 produces clinical improvement in people with these multisystem diseases.1,86,87 It has been inferred that B12 is acting as a potent nitric oxide scavenger and that this is the probable mode of action in the treatment of these multisystem diseases. 1,87 People with these diseases report essentially across-the-board improvement in symptoms when treated with hydroxocobalamin, suggesting that it acts to lower the basic etiologic mechanism of these diseases, consistent with a nitric–oxide scavenging mechanism.
In order to act as a nitric oxide scavenger, hydroxocobalamin and the chemically similar aquacobalamin must have the cobalt at the center of the molecule reduced from the cobalt III form to cobalt II.88 Such reduction is a process that occurs in vivo and is necessary for all cobalamins to have vitamin B12 activity as well as for hydroxocobalamin to serve as a nitric oxide scavenger.
Nitric oxide does not have a direct role in the central couplet, but it does serve as a direct precursor of peroxynitrite, such that nitric oxide scavenging will inevitably lower peroxynitrite levels in vivo. It can be argued, therefore, that hydroxocobalamin will act to lower the peroxynitrite end of the central couplet by scavenging nitric oxide.
Summary and Overall Strategy
Of the 10 agents/classes of agents described above that are known or predicted to lower the central couplet, nine individually appear to produce substantial improvements in this group of diseases, based on clinical trial studies, clinical observations, or both. The only one of the nine for which this is not true is oral ascorbate. These observations make the central couplet an attractive part of the cycle to focus on in trying to obtain substantial numbers of cures for these diseases. The question being raised here is whether combinations of these ten, especially combinations designed to effectively lower the central couplet, when added to the strategy that I previously advocated for treatment of these diseases, will produce such cures.
That strategy suggested here is as follows: Avoid stressors that will otherwise upregulate the NO/ONOO− cycle while using multiple agents that each lower one or more aspects of the cycle and collectively should lower several of its aspects.1-3 There are multiple approaches, each using such a multiple agent strategy, although the one that I have most worked on is the Allergy Research Group nutritional support protocol, which appears to produce positive responses in 80% to 85% of such patients. In general, such multiple agent approaches seem to have been effective in producing clinical improvements in most such patients but have failed to give any substantial numbers of cures, based on published information (thetenthparadigm.org/arg.htm).2,3
I think that the basic problem has been the failure to effectively downregulate the central couplet of the NO/ONOO− cycle. The proposal here is that we should add a second phase to these previous therapeutic approaches, one aimed at lowering that central couplet. More specifically, this means using agents that lower peroxynitrite and/or its breakdown products on the one hand; it also means using agents that increase BH4 availability on the other. Increased BH4 availability can be produced by using agents that reduce oxidized products of BH4 back to BH4. Such increased BH4 availability can also be produced by agents that induce the enzyme GTP cyclohydrolase I, the first and rate-limiting enzyme in the de novo pathway for the synthesis of BH4. What I have provided, then, is an overall strategy for getting some cures and a description of ten agents/classes of agents that should be useful in carrying out such a strategy. I have not, however, provided a detailed protocol for getting such cures.
I do think that it is possible that IV buffered ascorbate alone, when added to one of these broad-ranging protocols lowering the NO/ONOO− cycle and avoiding stressors that will raise the cycle, may be effective in obtaining some cures. I suspect, however, that most of the other agents that lower the central couplet should be used as multiagent combinations. And it is quite possible that even repeated IV ascorbate will be improved by using some of the other agents/classes of agents. The general strategy is to lower both ends of the couplet simultaneously, and probably repeatedly to progressively lower the cycle into insignificance. There is predicted to be synergistic interactions when using agents that work simultaneously to lower both ends of the central couplet.
I would be delighted to work with physicians and other health-care providers who are interested in exploring this approach.
If the view proposed in this article can be shown to be correct, then we will be in a new era in medicine. That will be true even if the relevance of this approach is limited to such diseases as CFS/ME, MCS, and FM. If other proposed NO/ONOO− cycle diseases, such as tinnitus, Parkinson's, Alzheimers, ALS, asthma, autism, and MS, can also be cured by this approach, then the impact on medicine will be comparable to the previous biggest therapeutic breakthrough, the development of wide-spectrum antibiotics.
Is this all delusional optimism? Clearly, we won't know until we look. But what we do know is that all of these diseases are chronic diseases, with cases of each apparently initiated by stressors that should be able to initiate the cycle. And we have evidence with all of them for important roles of such cycle elements as oxidative stress, inflammatory biochemistry, mitochondrial dysfunction, and excessive NMDA activity. Where they have been looked at, we also have evidence for BH4 depletion and NF-κB elevation. It is difficult to see how these cycle elements could be involved unless the NO/ONOO− cycle or something very similar to it is not central to the etiology of these diseases.
Mechanisms have consequences. It is time, in my view, for the sufferers of these diseases to fully benefit from the predictions of the NO/ONOO− cycle mechanism.
Martin L. Pall, PhD
Professor emeritus of biochemistry and basic medical sciences, Washington State University, and research director, the Tenth Paradigm Research Group
638 NE 41st Ave.
Portland, OR 97232-3312
USA
503-232-3883
martin_pall@wsu.edu
Notes
1. Pall ML. Explaining "Unexplained Illnesses": Disease Paradigm for Chronic Fatigue Syndrome, Multiple Chemical Sensitivity, Fibromyalgia, Post-Traumatic Stress Disorder, Gulf War Syndrome and Others. Harrington Park Press; 2007.
2. Pall ML. Multiple chemical sensitivity: toxicological questions and mechanisms. In: Ballantyne B, Marrs TC, Syversen T, eds. General and Applied Toxicology. 3rd ed. London: John Wiley & Sons; 2009:2303–2352.
3. Pall ML. The NO/ONOO− cycle mechanism as the cause of chronic fatigue syndrome/myalgia encephalomyelitis. Chapter 2 in: Svoboda E, Zelenjcik K, eds. Chronic Fatigue Syndrome: Symptoms, Causes and Prevention. Nova Publishers; 2009.
4. Pall ML. The NO/ONOO− cycle as the cause of fibromyalgia and related illnesses: etiology, explanation and effective therapy. Chapter 2 in: Pederson JA, ed. New Research in Fibromyalgia. Nova Science Publishers; 2006:39–59.
5. Pall ML. Elevated, sustained peroxynitrite levels as the cause of chronic fatigue syndrome. Med Hypotheses. 2000:54:115–125.
6. Pall ML. Common etiology of posttraumatic stress disorder, fibromyalgia, chronic fatigue syndrome and multiple chemical sensitivity via elevated nitric oxide/peroxynitrite. Med Hypotheses. 2001;57:139–145.
7. Pall ML, Satterlee JD. Elevated nitric oxide/peroxynitrite mechanism for the common etiology of multiple chemical sensitivity, chronic fatigue syndrome and posttraumatic stress disorder. Ann NY Acad Sci. 2001;933:323–329.
8. Hinoi E, Takarada T, Ueshima T, Tsuchihashi Y, Yoneda Y. Glutamate signaling in peripheral tissues. Eur J Biochem. 2004:271:1–13.
9. Milstien S, Katusic Z. Oxidation of tetrahydrobiopterin by peroxynitrite: Implications for vascular endothelial function. Biochem Biophys Res Commun. 1999;263:681–684.
10. Laursen JB, Somers M, Kurz S, et al. Endothelial regulation of vasomotion in apoE-deficient mice: implications for interactions between peroxynitrite and tetrahydrobiopterin. Circulation. 2001;103:1282–1288.
11. Pall ML. Nitric oxide synthase partial uncoupling as a key switching mechanism for the NO/ONOO− cycle. Med Hypotheses. 2007;69:821–825.
12. Foxton RH, Land JM, Heales SJ. Tetrahydrobiopterin availability in Parkinson's and Alzheimer's disease; potential pathogenic mechanisms. Neurochem Res. 2007;32:751–756.
13. Pall ML. Do sauna therapy and exercise act by raising the availability of tetrahydrobiopterin? Med Hypotheses. 2009;73:610–613.
14. Teitelbaum JE, Bird B, Greenfield, et al. Effective treatment of chronic fatigue syndrome and fibromyalgia: arandomized, double-blind, placebo-controlled, intent to treat study. J Chronic Fatigue Syndr. 2002;8(2):3–28.
15. Pall ML, Bedient SA. The NO/ONOO− cycle as the etiological mechanism of tinnitus. Int Tinnitus J. 2007;13:99–104.
16. Pall ML. Post-radiation syndrome as a NO/ONOO− cycle, chronic fatigue syndrome-like disease. Med Hypotheses. 2008;71:537–541.
17. Griendling KK, Ushio-Fukai. Reactive oxygen species as mediators of angiotensin II signalling. Regul Pept. 2000;91:21–27.
18. Landmesser U, Dikalov S, Price SR, et al. Oxidation of tetrahydrobiopterin leads to uncoupling of endothelial cell nitric oxide synthase in hypertension. J Clin Invest. 2003 Apr;111(8):1201–1209.
19. Schulz E, Jansen T, Wenzel P, Daiber A, Münzel T. Nitric oxide, tetrahydrobiopterin, oxidative stress, and endothelial dysfunction in hypertension. Antioxid Redox Signal. 2008;10:1115–1126.
20. Moens AL, Kass DA. Tetrahydrobiopterin and cardiovascular disease. Arterioscler Thromb Vasc Biol. 2006;26:2439–2244.
21. Padayatty SJ, Levine M. New insights into the physiology and pharmacology of vitamin C. CMAJ. 2001;164:353–355.
22. Padayatty SJ, Sun H, Wang Y, et al. Vitamin C pharmacokinetics: implications for oral and intravenous use. Ann Intern Med. 2004;140:533–537.
23. Levine M, Rumsey SC, Daruwala R, Park JB, Wang Y. Criteria and recommendations for vitamin C intake. JAMA. 1999;281:1415–1423.
24. Riordan HD, Hunninghake RB, Riordan NH, et al. Intravenous ascorbic acid: protocol for its application and use. P R Health Sci J. 2003;22:287–290.
25. Heuser G, Vojdani A. Enhancement of natural killer cell activity and T and B cell function by buffered vitamin C in patients exposed to toxic chemicals: the role of protein kinase C. Immunopharmacol Immunotoxicol. 1997;19:291–312.
26. Kodama M, Kodama T, Murakami M. The value of the dehydroepiandrosterone-annexed vitamin C infusion treatment in the clinical control of chronic fatigue syndrome (CFS). I. A Pilot study of the new vitamin C infusion treatment with a volunteer CFS patient. In Vivo. 1996;10:575–584.
27. Kodama M, Kodama T, Murakami M. The value of the dehydroepiandrosterone-annexed vitamin C infusion treatment in the clinical control of chronic fatigue syndrome (CFS). II. Characterization of CFS patients with special reference to their response to a new vitamin C infusion treatment. In Vivo. 1996;10:585–596.
28. Kodama M, Kodama T. Four problems with the clinical control of interstitial pneumonia, or chronic fatigue syndrome, using the megadose vitamin C infusion system with dehydroepiandrosterone-cortisol annex. In Vivo. 2006;20:285–291.
29. Levy, TE. Curing the Incurable: Vitamin C, Infectious Disease and Toxins. 3rd ed. Henderson, NV: LivOn Books; 2009.
30. Kuzkaya N, Weissmann N, Harrison DG, Dikalov S. Interactions of peroxynitrite, tetrahydrobiopterin, ascorbic acid and thiols: implications for uncoupling of endothelial nitric oxide synthase. J Biol Chem. 2003;278:22546–22554.
31. Whiteman M, Ketsawatsakul U, Halliwell B. A reassessment of the peroxynitrite scavenging activity of uric acid. Ann N Y Acad Sci. 2002;962:242–259. BH4 oxidation by carbonate and NO2 radical (cite also for ascorbate reduction)
32. Patel KB, Stratford MRL, Wardman P, Everett SA. Oxidation of tetrahydrobiopterin by biological radicals and scavenging of trihydrobiopterin radical by ascorbate. Free Rad Biol Med. 2002;32:203–211.
33. Hydrogen peroxide. See Padayatty et al.; Levine et al.; Riordan et al.; and Chen Q, Espey MG, Krishna MC, et al. Pharmacologic ascorbic acid concentrations selectively kill cancer cells: action as a pro-drug to deliver hydrogen peroxide to tissues. Proc Natl Acad Sci U S A. 2005;102,13604–13609.
34. Clément MV, Ramalingam J, Long LH, Halliwell B. The in vitro cytotoxicity of ascorbate depends on the culture medium used to perform the assay and involves hydrogen peroxide. Antioxid Redox Signal. 2001;3:157–163.
35. Shimizu S, Shiota K, Yamamoto S, Miyasaka Y, Ishii M, et al. Hydrogen peroxide stimulates tetrahydrobiopterin synthesis through the induction of GTP-cyclohydrolase I and increases nitric oxide synthase activity in vascular endothelial cells. Free Radic Biol Med. 2003;34:1343–1352.
36. Ishii M, Shimizu S, Wajima T, et al. Reduction of GTP cyclohydrolase I feedback regulating protein expression by hydrogen peroxide in vascular endothelial cells. J Pharmacol Sci. 2005;97:299–302.
37. Kalivendi S, Hatakeyama K, Whitsett J, Konorev E, Kalyanaraman B, Vásquez-Vivar J. Changes in tetrahydrobiopterin levels in endothelial cells and adult cardiomyocytes induced by LPS and hydrogen peroxide – a role for GFRP? Free Radic Biol Med. 2005;38,481–491.
38. Rees DC, Kelsey H, Richards JD. Acute haemolysis induced by high dose ascorbic acid in glucose-6-phosphate dehydrogenase deficiency. BMJ. 1993;306:841–842.
39. May JM. How does ascorbic acid prevent endothelial dysfunction? Free Radic Biol Med. 2000;28:1421–1429.
40. Fotherby MD, Williams JC, Forster LA, Craner P, Ferns GA. Effect of vitamin C on ambulatory blood pressure and plasma lipids in older persons. J Hypertens. 2000;18:411–415.
41. Bates CJ, Walmsley CM, Prentice A, Finch S. Does vitamin C reduce blood pressure? Results of a large study of people aged 65 or older. J Hypertens. 1998;16:925–932.
42. Gibson PR, Elms AN-M, Ruding LA. Perceived treatment efficacy for conventional and alternative therapies reported by persons with multiple chemical sensitivity. Environ Health Perspect. 2003;111:1498–1504.
43. Rea WJ. Chemical Sensitivity. Boca Raton, FL: Lewis Publishers;1992.
44. Crinnion W. Components of practical clinical detox programs – sauna as a therapeutic tool. Altern Ther Health Med. 2007;13:S154–S156. Baird DN, Rea WJ. The temperomandibular joint implant controversy. Part II: its clinical implications. J Nutr Environ Med. 1999;9:209–222.
45. Rogers SA. Detoxify or Die. Syracuse, NY: Prestige Publishers; 2002.
46. Krop J. Chemical sensitivity after intoxication at work with solvents: response to sauna therapy. J Altern Complement Med. 1998;4:77–86.
47. Masuda A, Munemoto T, Tei C. A new treatment: thermal therapy for chronic fatigue syndrome. Nippon Rinsho. 2007;65:1093–1098.
48. Brockow T, Wagner A, Franke A, Offenbacher M, Resch KL. A randomized controlled trial on the effectiveness of mild water-filtered near infrared whole-body hyperthemia as an adjunct to standard multimodal rehabilitation in the treatment of fibromyalgia. Clin J Pain. 2007;23:67–75.
49. Matsushita K, Masuda A, Tei C. Efficacy of Waon therapy for fibromyalgia. Intern Med. 2008;47:1473–1476.
50. Pall ML. Do sauna therapy and exercise act by raising the availability of tetrahydrobiopterin? Med Hypotheses. 2009;73:610–613.
51. Teitelbaum JE, Johnson C, St Cyr J. The use of D-ribose in chronic fatigue syndrome and fibromyalgia: a pilot study. J Altern Complement Med. 2006;12:857–862.
52. Ducrocq C, Blanchard B, Pignatelli B, Ohshima H. Peroxynitrite: an endogenous oxidizing and nitrating agent. Cell Mol Life Sci. 1999;55:1068–1077.
53. Geren W, Bendich A, Bodansky O, Brown GB. The fate of uric acid in man. J Biol Chem. 1950;183:21–31.
54. Wei Y, Chen L, Chen J, Ge L, He RQ. Rapid glycation with D-ribose induces globular amyloid-like aggregations of BSA with high cytotoxicity to SH-SY5Y cells. BMC Cell Biol. 2009 Feb 13;10:10.
55. Bailey AJ, Sims TJ, Avery NC, Halligan EP. Non-enzymic glycation of fibrous collagen: reaction products of glucose and ribose. Biochem J. 1995;305( Pt 2):385–390.
56. Yan H, Harding JJ. Glycation-induced inactivation and loss of antigenicity of catalase and superoxide dismutase. Biochem J. 1997;328(Pt 2):599–605.
57. Haskó G, Sitkovsky MV, Szabó C. Immunomodulatory and neuroprotective effects of inosine. Trends Pharmacol Sci. 2004;25:152–157.
58. Tilley SL, Wagoner VA, Salvatore CA, Jacobson MA, Koller BH. Adenosine and inosine increase cutaneous vasopermeability by activating A(3) receptors on mast cells. J Clin Invest. 2000;105:361–367.
59. Hyndman ME, Verma S, Rosenfeld RJ, Anderson TJ, Parsons HG. Interaction of 5-methyltetrahydrofolate and tetrahydrobiopterin on endothelial function. Am J Physiol Heart Circ Physiol. 2002 Jun;282(6):H2167–172.
60. Moat SJ, Clarke ZL, Madhavan AK, Lewis MJ, Lang D. Folic acid reverses endothelial dysfunction induced by inhibition of tetrahydrobiopterin biosynthesis. Eur J Pharmacol. 2006 Jan 20;530(3):250–258.
61. Verhaar MC, Wever RM, Kastelein JJ, van Dam T, Koomans HA, Rabelink TJ. 5-methyltetrahydrofolate, the active form of folic acid, restores endothelial function in familial hypercholesterolemia. Circulation. 1998 Jan 27;97(3):237–241.
62. Gori T, Burstein JM, Ahmed S, Miner SE, Al-Hesayen A, Kelly S, Parker JD. Folic acid prevents nitroglycerin-induced nitric oxide synthase dysfunction and nitrate tolerance: a human in vivo study. Circulation. 2001 Sep 4;104(10):1119–1123.
63. Antoniades C, Shirodaria C, Warrick N, et al. 5-methyltetrahydrofolate rapidly improves endothelial function and decreases superoxide production in human vessels: effects on vascular tetrahydrobiopterin availability and endothelial nitric oxide synthase coupling. Circulation. 2006;114:1193–1201.
64. Rezk BM, Haenen GRMM, van der Vijgh WJF, Bast A. Tetrahydrofolate and 5-methyltetrahydrofolate are folates with high antioxidant activity. Identification of the antioxidant pharmacophore. FEBS Lett. 2003;555:601–605
65. Offer T, Ames BN, Bailey SW, Sabens EA, Nozawa M, Ayling JE. 5-Methyltetrahydrofolate inhibits photosensitization reactions and strand breaks in DNA. FASEB J. 2007;21:2101–2107.
66. Doshi SN, McDowell IF, Moat SJ, et al. Folic acid improves endothelial function in coronary artery disease via mechanisms largely independent of homocysteine lowering. Circulation. 2002;105:22–26.
67. Jacobson W, Saich T, Borysiewicz LK, Behan WM, Behan PO, Wreghitt TG. Serum folate and chronic fatigue syndrome. Neurology. 1993;43:2645–2647.
68. Gerwin RD. The management of myofascial pain sydromes. J Musculoskeletal Pain. 1993;1(3/4):83–94.
69. Lundell K, Qazi S, Eddy L, Uckun FM. Clinical activity of folinic acid in patients with chronic fatigue syndrome. Arzneimittelforschung. 2006;56:399–404.
70. Danfors T, von Knorring AL, Hartvig P, et al. Tetrahydrobiopterin in the treatment of children with autistic disorder: a double-blind placebo-controlled crossover study. J Clin Psychopharmacol. 2005;25:485–489.
71. Fernell E, Watanabe Y, Adolfsson I, et al. Possible effects of tetrahydrobiopterin treatment in six children with autism – clinical and positron emission tomography data: a pilot study. Dev Med Child Neurol. 1997;39:313–318.
72. Naruse H, Hayashi T, Takesada M, Nakane A, Yamazaki K. Metabolic changes in aromatic amino acids and monoamines in infantile autism and development of new treatment related to the finding. [In Japanese.] No To Hattatsu. 1989 Mar;21:181–189.
73. Choi HJ, Lee SY, Cho Y, No H, Kim SW, Hwang O. Tetrahydrobiopterin causes mitochondrial dysfunction in dopaminergic cells: implications for Parkinson's disease. Neurochem Int. 2006;48:255–262.
74. Lee SY, Moon Y, Hee Choi D, Jin Choi H, Hwang O. Particular vulnerability of rat mesencephalic dopaminergic neurons to tetrahydrobiopterin: Relevance to Parkinson's disease. Neurobiol Dis. 2007;25:112–120.
75. Anastasiadis PZ, Bezin L, Gordon LJ, Imerman B, Blitz J, Kuhn DM, Levine RA. Vasoactive intestinal peptide induces both tyrosine hydroxylase activity and tetrahydrobiopterin biosynthesis in PC12 cells. Neuroscience. 1998;86:179–189.
76. McCarty MF. Scavenging of peroxynitrite-derived radicals by flavonoids may support endothelial NO synthase activity, contributing to the vascular protection associated with high fruit and vegetable intakes. Med Hypoth. 2008;70:170–181.
77. Ippoushi K, Takeuchi A, Azuma K. Prevention of peroxynitrite-induced oxidation and nitration reactions by ellagic acid. Food Chem. 2008;112:185–188.
78. Basu A, Penugonda K. Pomegranate juice: a heart-healthy fruit juice. Nutr Rev. 2009;67:49–56.
79. Perez-Vizcaino F, Duarte J, Andriantsitohaina R. Endothelial function and cardiovascular disease: effects of quercetin and wine polyphenols. Free Radic Res. 2006;40:1054–1065.
80. Ghosh D, Scheepens A. Vascular action of polyphenols. Mol Nutr Food Res. 2009;53:322–331.
81. Schmitt CA, Dirsch VM. Modulation of endothelial nitric oxide by plant-derived products. Nitric Oxide. 2009;21:77–91.
82. Romero M, Jimenez R, Sanchez M, et al. Quercetin inhibits vascular superoxide production induced by indothelin-1: Role of NADPH oxidase, uncoupled eNOS and PKC. Atherosclerosis. 2009;202:58–67.
83. Ellis FR, Nasser S. A pilot study of vitamin B12 in the treatment of tiredness. Br J Nutr 1973;30:277–283; Lawhorne L, Ringdahl D. Cyanocobalamin injections for patients without documented deficiency. JAMA. 1989;261:1920–1923.
84. Kaslow JE, Rucker L, Onishi R. Liver extract-folic acid-cyanocobalamin vs placebo for chronic fatigue syndrome. Arch Intern Med. 1989;149:2501–2503.
85. Lapp CW. Using vitamin B-12 for management of CFS. CFIDS Chron. 1999;12(6):14–16.
86. Hoh D. Treatment of the Cheney clinic. CFIDS Chron. 1998;1(4):13–14.
87. Pall ML. Cobalamin used in chronic fatigue syndrome therapy is a nitric oxide scavenger. J Chronic Fatigue Syndr. 2001;8(2):39–44.
88. Wolak M, Zahl A, Schneppensieper T, Stochel G, van Eldik R. Kinetics and mechanism of the reversible binding of nitric oxide to reduced cobalamin B(12r) (Cob(II)alamin). J Am Chem Soc. 2001;123:9780–8791.
The NO/ONOO− cycle is named for two of its many elements, nitric oxide (NO) and peroxynitrite (ONOO−) and is pronounced "no, oh no."
S-adenosylmethionine (SAMe) is the main direct methyl donor in living organisms, being produced by the methylation cycle and acting in turn to methylate many different substrates in the cell. There have been many claims that these illnesses are caused by lowered methylation cycle activity. I think that these claims not valid. There is a modest lowering of methylation activity caused by peroxynitrite-mediated 5-MTHF oxidation, but whether such modest lowering of methylation has any causal role is unclear. What should be clear is that such a modest methylation cycle lowering should be normalized by an effective downregulation of the NO/ONOO− cycle, including especially the central couplet. That is the treatment approach explored in this article is the approach that should be used to normalize various properties of these NO/ONOO− cycle diseases, including the modest lowering of methylation cycle activity.
![]()
Consult your doctor before using any of the treatments found within this site.
![]()
Subscriptions are available for Townsend Letter, the Examiner of Alternative Medicine magazine, which is published 10 times each year.
Search our pre-2001
archives for further information. Older issues of the printed magazine
are also indexed for your convenience.
1983-2001
indices ; recent indices
Once you find the magazines you'd like to order, please use our convenient form, e-mail subscriptions@townsendletter.com, or call 360.385.6021 (PST).
Who are we? | New
articles | Featured topics |
Tables of contents | Subscriptions | Contact
us | Links | Classifieds | Advertise | Alternative
Medicine Conference Calendar | Search site | Archives |
EDTA
Chelation Therapy | Home
© 1983-2010 Townsend Letter for Doctors
& Patients
All rights reserved.
Website by Sandy Hershelman
Designs
March 30, 2010

![]()