|
Page 1, 2
An attack is proof that one is out of control. Never run away from any kind of challenge, but do not try to suppress or control an opponent unnaturally. Let attackers come any way they like and then blend with them. … Redirect each attack and get firmly behind it.
– Morihei Ueshiba, founder of the martial art Aikido
The conventional model of cancer is dominated by the idea that malignant cells are aberrations, or good cells that suffer various genetic mutations and turn against their host. It is a model of cancer that justifies not only the conventional approach to cancer treatment – namely, to directly kill as many malignant cells as possible – but also underlies many of the alternative therapies in common use. It is a model that, with a few noteworthy exceptions, has been overwhelmingly underwhelming in its achievements.
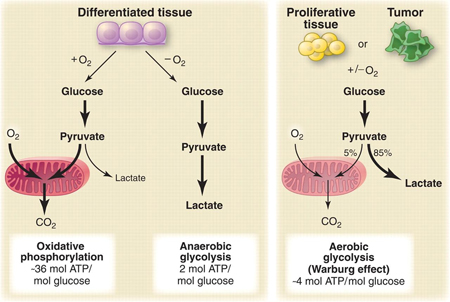
A different model for understanding the malignant process was famously proposed by Dr. Otto Warburg in 1931.Warburg was awarded the Nobel Prize over 80 years ago for his discovery that cancer cells use glucose at a rate 10 to 50 times higher than healthy cells through direct glycolysis, an anaerobic pathway. He observed that a characteristic of malignant cells is their continued reliance on glycolysis even when the oxygen supply is restored to the cell, a condition now known as the Warburg effect. His seminal work on the role of hypoxia in cancer development continues to influence researchers to this day. This model takes a radical and provocative approach to cancer: the transformation of the cell from healthy to malignant is an adaptation.
In this model, healthy cells become exposed to some set of circumstances that leave it two options: adapt to these circumstances, or die. Many cells will, in fact, die, leading to various types of pathology (lung fibrosis, liver sclerosis/cirrhosis, neuronal death leading to Parkinson's, and many others). Whether it is by necrosis or apoptosis, cells unable to adapt must meet an early death.
However, when those potentially lethal circumstances are fundamentally linked to reduced oxygen availability due to toxic exposure, impaired circulation, chronic inflammation, or many other causes, cells have the potential to adapt. They rewire themselves, genetically and metabolically, to survive in spite of their circumstances. Enzymes upregulate and downregulate, embryonic genes activate, some metabolic pathways get shut down, while others light up, and so on.1-3 The malignant cell represents an exquisitely coordinated and nonrandom set of transformations that lead to cell survival in spite of its circumstances.4,5
It is within this context that the model of cancer as a metabolic disease gains its purchase. In this model the cancer cell is no longer an adversary to kill, but an adaptation that is no longer needed. Warburg demonstrated that there is potentially a point of no return: after some time in that adapted state, the cell isn't likely to revert to normalcy.
At the same time, Ueshiba reminds us that to control our challenger, our best move is to "redirect … and get firmly behind it." This is the foundation of the metabolic approach to cancer treatment. Building a rational metabolic approach to cancer starts with an understanding of cancer cell metabolism. Here are the basics.
Metabolic Factors
Stem cells are progenitor cells that endlessly give rise to the new cells that regenerate our bodies, beginning to end. In healthy tissue, stem cells exist with relatively few mitochondria, and sit in a hypoxic "niche." They give rise, upon signaling, to "daughter" cells that migrate out of that niche to become differentiated cells of that tissue. These daughter cells move through an increasing oxygen gradient, and in the process they lose the characteristics of the parent stem cell, such as endless replicative potential and lack of differentiation.
If local hypoxia has reduced or abolished this oxygen gradient moving away from the niche, or if tissue damage has impaired appropriate tissue-based differentiation signals to the migrating cell, the daughter cell retains its undifferentiated state and potentially its replicative potential. It also downregulates its gap-junction communication, leading to dissociation from adjacent cells and tissue, and characteristic independent behavior.6
These events are often accompanied by mutations in regulatory genes such as RAS, TP53, MYC, Oct-4, and others. Whether mutations are primary or secondary to malignancy is unclear. Nevertheless, once in place, genetic mutations unquestionably drive the malignant phenotype.
Another effect of the malignant rewiring of the cell has to do with its ability to quench the relatively high production of reactive oxygen species (ROS) generated within malignant cells. For example, upregulation of the pentose phosphate pathway (PPP) leads to generation of the ROS-quenching antioxidant glutathione. This pathway also generates the ribose required for building the DNA needed to fill this burgeoning population of malignant cells.
A central feature of the malignant cell phenotype is its rewiring of the dominant energy pathway within the cell. Healthy cells use oxidative phosphorylation via the Krebs cycle to generate the bulk of its ATP energy. Cancer cells, though, generally show a dominance of anaerobic glycolysis, whereby the pyruvate generated from glucose is diverted – via the upregulation of several enzymes including PKM2 and LDH-A and downregulation of pyruvate dehydrogenase (PDH) – into NAD and lactic acid production.
These are profound shifts that supply the high levels of fatty acids and ribose needed to sustain the exponential rate of cell division. While glycolysis is far less efficient in producing ATP, the shift represents an adaptation to the growth and reproduction needs of the cell. As mentioned above, cancer cells consume 10 to 50 times as much glucose as healthy cells to sustain this seemingly inefficient metabolism, and they do this for at least 3 reasons:
- While aerobic glycolysis produces less ATP per molecule of glucose, cancer cells can produce ATP this way much faster, with estimates of up to 100 times faster than normal cells. Speed trumps efficiency.
- Cancer cells using aerobic glycolysis produce many biosynthetic precursors such as ribulose-5-p which are the building blocks for the production of proteins, lipids, and DNA required by the rapidly dividing cancer cells.
- Signals for apoptosis – programmed cell death – are largely buried within the cell's mitochondria. By routing energy pathways away from mitochondria, malignant cells avoid tripping these switches, thus preserving their immortal status.
It should be emphasized once again that, contrary to popular portrayal of these events, the genetic changes driving these adaptations are not random. Cancer cells pursue each of these adaptations for survival in a concerted and coordinated way.
Apoptotic Factors
Apoptosis is a programmed cell suicide in response to external or internal stimuli. For many years the mechanisms behind apoptosis were poorly understood. More recent probing into cells has elucidated a great many of the details behind the process.
The first pathway for this process is the extrinsic, or the cytoplasmic, pathway. This is carried out by a member of the tumor necrosis factor family appropriately called the Fas death receptor. The Fas receptor is located on the surface of the cell and is triggered by a ligand, which is a molecule that when produced fits into a complex receptor site on the Fas protein, like a key fits into a lock. When the ligand is a perfect fit for that site it produces the FADD (Fas-associated death domain) protein, along with caspase 8 and caspase 10. These proteins then trigger a cascade of additional caspases which essentially dismantle the cell by cleaving its internal proteins as well as its protein scaffolding.
The second pathway is the intrinsic or mitochondrial pathway mentioned previously in this article. When this pathway is stimulated it triggers a release of cytochrome-c from the mitochondria which activates the apoptotic death signal. Cytochrome-c is a heme protein that is bound to the inner wall of the mitochondrial membrane. When cytochrome-c is released it triggers a release of capase-9, which then releases caspase-3 and caspase-7. Metabolically rewiring the cell away from use of the mitochondria avoids activation of these caspases, an essential step in avoiding apoptosis and maintaining the malignant phenotype.
It is easy to see that both the extrinsic (cytoplasmic) and the intrinsic (mitochondrial) apoptotic pathways converge upon the formation of caspases. That is, all roads lead to caspase-mediated apoptosis. These caspases are in a class of molecules called proteases and their job is to cleave structural and regulatory proteins of cells determined to be a threat. It is interesting to note that, as with so much in physiology, the two pathways balance each other: overexpression of one path can inhibit the other and vice versa. Even malignant cells have self-regulatory mechanisms in place, which is of course no accident.
Clinical Relevance with Attention to Fermented Wheat Germ Extract
In my practice, I see patients with all types and stages of cancer, creating treatment plans rooted in this belief that cancer is fundamentally not an aberration, but a metabolic adaptation. I'm utilizing a number of natural therapies, and often bring in conventional medications as well, always with an eye toward low toxicity and least potential to cause harm. Some of these metabolic therapies include alpha-lipoic acid, dichloroacetate (DCA), organic germanium, low-dose naltrexone, butyrate, high-dose pancreatic enzymes, and many others.
Recently, I have become very interested in fermented wheat germ extract, not only due to its successful clinical studies against melanoma, colorectal cancer, oral cancer, and others, but also because its mechanism of action is very much in alignment with the approach to cancer as a metabolic disorder.
To review, fermented wheat germ extract (FWGE) is made through a fermentation process whereby wheat germ and baker's yeast are combined with water, filtered, dried, and packaged for consumption. Nobel laureate Dr. Albert Szent-Györgyi initially proposed the use of FWGE as an anticancer agent. He hypothesized that disorders of metabolism might play important roles in cancer development, and found that high redox potential quinones such as those naturally occurring in wheat germ could block cell replication. He suggested that they might prove useful in reversing disorders of cellular metabolism.7
Promotion of Apoptosis
Early research on FWGE found that it influences apoptosis via several molecular pathways. First, FWGE increases levels of cytochrome-c. As discussed previously, cytochrome-c is a very important member of the mitochondrial apoptotic pathway and the presence of cytochrome-c in the cytosol is a clear marker of mitochondria-induced apoptosis. Next, FWGE effects apoptosis indirectly through its cleavage of PARP, or poly ADP ribose polymerase. PARP is a family of proteins involved in a number of cellular processes primarily involving DNA repair and programmed cell death (i.e., if cells don't repair their DNA with PARP, they die).
Malignant cells utilize PARP to repair DNA damage and prevent activation of the mitochondrial apoptosis pathway whereby phosphatidylserine is shuttled from the inner to the outer membrane leaflet, a shuttle activated by caspase proteases, and most specifically caspase-3. FWGE activates caspase-3 and a number of other apoptosis-associated enzymes, including Bax and Bcl-2, and CyclinD1. The result is cleavage of PARP. This allows for both the apoptotic externalization of phosphatidylserine as well as prevention of DNA repair by PARP specifically in cancer cells.
Page 1, 2
|




